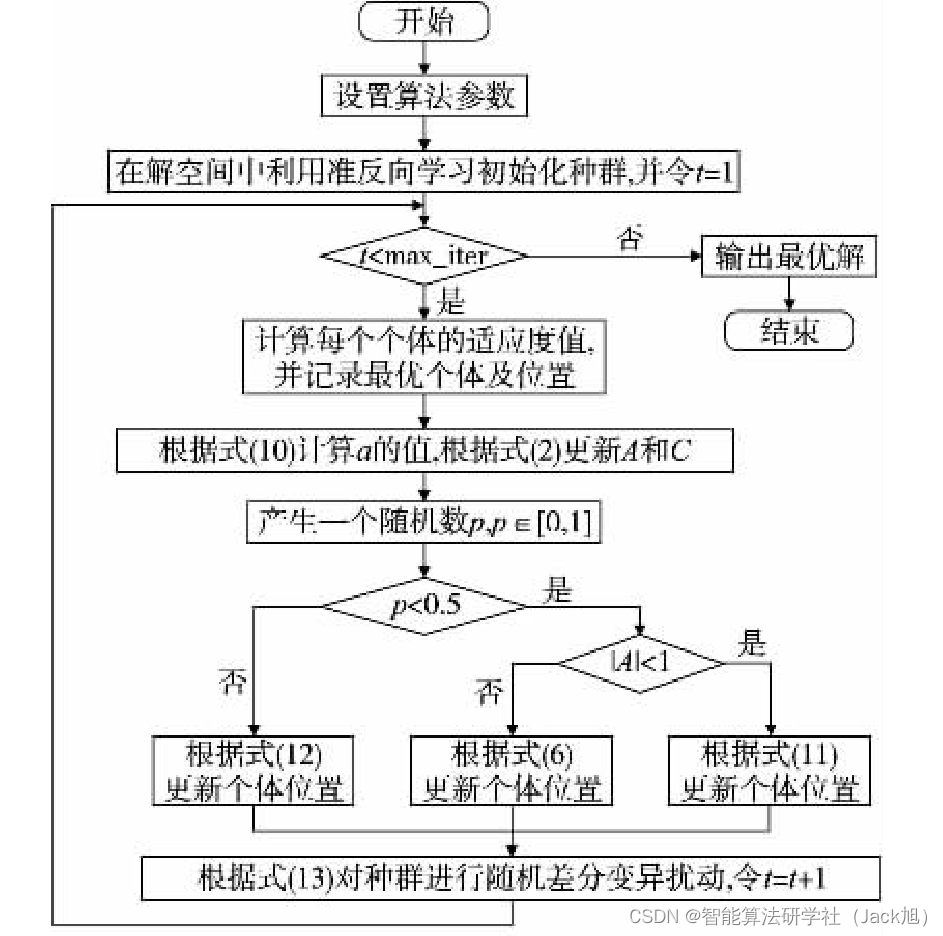
Gondro 等人
[
8
]
{ }^{[8]}
[8] 指出高质量的初始化种群对算法的求解精 度和收敛速度等性能有很大的帮助。然而, 基本的鲸鱼优化算 法采用的是随机初始化的方法, 该方法随机性很大, 不能保证 初始种群的多样性。为了保证初始化种群的多样性, 采用了反 向学习的改进方法, 即准反向学习来初始化种群。 反向学习
[
6
,
9
,
10
]
{ }^{[6,9,10]}
[6,9,10] 是在明确变量的范围边界, 通过按照一定 的规则来求其对应的反向解, 其详细内容如下: 假设鲸鱼种群的规模为
N
N
N, 搜索空间为
d
d
d 维, 第
i
i
i 只鲸鱼 在第
d
d
d 维空间中的位置可以表示为
X
i
=
(
x
i
1
,
x
i
2
,
⋯
,
x
i
d
)
(
i
=
1
\boldsymbol{X}_{i}=\left(x_{i}^{1}, x_{i}^{2}, \cdots, x_{i}^{d}\right)(i=1
Xi=(xi1,xi2,⋯,xid)(i=1,
2
,
⋯
,
N
)
,
x
i
j
∈
[
a
i
j
,
b
i
j
]
(
j
=
1
,
2
,
⋯
,
d
)
,
a
i
j
2, \cdots, N), x_{i}^{j} \in\left[a_{i}^{j}, b_{i}^{j}\right](j=1,2, \cdots, d), a_{i}^{j}
2,⋯,N),xij∈[aij,bij](j=1,2,⋯,d),aij 和
b
i
j
b_{i}^{j}
bij 分别表示
x
i
j
x_{i}^{j}
xij 的 下界和上界,其对应反向解如下:
x
^
i
j
=
a
i
j
+
b
i
j
−
x
i
j
(7)
\hat{x}_{i}^{j}=a_{i}^{j}+b_{i}^{j}-x_{i}^{j} \tag{7}
x^ij=aij+bij−xij(7) 在实际应用中, 反向解的效果并不一定很好, 本文为了解 决这一问题采用了准反向解的方法, 该方法可以根据式 (7) 演 变得到:
x
i
j
^
=
{
rand
(
avg
i
j
,
x
^
i
j
)
x
i
j
⩽
avg
i
j
rand
(
x
^
i
j
,
avg
i
j
)
x
i
j
>
avg
i
j
(8)
\hat{x_{i}^{j}}= \begin{cases}\operatorname{rand}\left(\operatorname{avg}_{i}^{j}, \hat{x}_{i}^{j}\right) & x_{i}^{j} \leqslant \operatorname{avg}_{i}^{j} \\ \operatorname{rand}\left(\hat{x}_{i}^{j}, \operatorname{avg}_{i}^{j}\right) & x_{i}^{j}>\operatorname{avg}_{i}^{j}\end{cases} \tag{8}
xij^=⎩⎨⎧rand(avgij,x^ij)rand(x^ij,avgij)xij⩽avgijxij>avgij(8) 其中:
avg
i
j
=
b
i
j
−
a
i
j
2
;
(
a
,
b
)
\operatorname{avg}_{i}^{j}=\frac{b_{i}^{j}-a_{i}^{j}}{2} ;(a, b)
avgij=2bij−aij;(a,b) 代表在
a
、
b
a 、 b
a、b 之间的随机数。 随机产生的
N
N
N 个初始个体和准反向学习
[
9
]
{ }^{[9]}
[9] 求得的
N
N
N 个准 反向解将其合并, 然后通过一种种群多样性最大化的篮选机 制, 从这
2
N
2 N
2N 个个体中选择种群多样性最大化的
N
N
N 个个体。 为了使种群获得比较好的初始化种群, 本文将随机解与其 对应的准反向解进行取优化处理, 这样既保证了种群的多样性, 同时又能使种群较快地收敛到全局最优解。其数学模型如下:
fit
(
X
)
>
fit
(
V
∨
)
?
X
,
X
V
(9)
\text { fit }(X)>\text { fit }(\stackrel{\vee}{V}) ? X, \stackrel{V}{X} \tag{9}
fit (X)> fit (V∨)?X,XV(9) 其中: fit 为适应度函数;
X
X
X 和
X
X
X 分别表示随机产生的个体和准 反向学习产生的个体。
2.2 非线性收敛因子
与其他群体智能优化算法类似, 鲸鱼优化算法在寻优过程 中同样会遇到全局搜索能力和局部开发能力不平衡的现象。 在基本鲸鱼优化算法中, 其掌握鲸鱼进行全局搜索还是局部搜 索的是参数
∣
A
∣
|A|
∣A∣, 当参数
∣
A
∣
⩾
1
|A| \geqslant 1
∣A∣⩾1 时, 算法以
0.5
0.5
0.5 的概率进行随 机全局搜索, 当
∣
A
∣
<
1
|A|<1
∣A∣<1 时, 算法进行局部开发。由于收玫因子
a
a
a 进行线性变化并不能很好地调节全局搜索能力和局部开发 能力, 因此本文提出来一种非线性收敛因子
[
5
,
11
,
12
]
{ }^{[5,11,12]}
[5,11,12] 为
a
=
2
−
2
sin
(
μ
t
m
a
x
i
t
e
r
π
+
φ
)
(10)
a=2-2 \sin \left(\mu \frac{t}{{ max_iter }} \pi+\varphi\right) \tag{10}
a=2−2sin(μmaxitertπ+φ)(10) 其中: max_iter 为最大迭代次数;
t
t
t 为当前迭代次数;
μ
\mu
μ 和
φ
\varphi
φ 是其 表达式相关参数, 选取
μ
=
1
2
,
φ
=
0
\mu=\frac{1}{2}, \varphi=0
μ=21,φ=0 。
2.3 自适应权重策略与随机差分法变异策略
鲸鱼优化算法在后期局部开发时易陷入局部最优, 出现早 熟收敛的现象, 为了使算法能够保持种群的多样性并且能够及 时跳出局部最优, 提出来一种自适应权重策略和随机差分变异策略 自适应权重策略数学表达式如下:
ω
=
1
−
e
t
max
iter
−
1
e
−
1
,
X
(
t
+
1
)
=
ω
⋅
X
p
(
t
)
−
A
⋅
D
(11)
\omega=1-\frac{e^{\frac{t}{\max \text { iter }}-1}}{e-1}, \boldsymbol{X}(t+1)=\omega \cdot \boldsymbol{X}_{p}(t)-\boldsymbol{A} \cdot \boldsymbol{D} \tag{11}
ω=1−e−1emax iter t−1,X(t+1)=ω⋅Xp(t)−A⋅D(11)
X
(
t
+
1
)
=
ω
⋅
X
p
(
t
)
+
D
⋅
e
b
l
⋅
cos
(
2
π
l
)
(12)
\boldsymbol{X}(t+1)=\omega \cdot \boldsymbol{X}_{p}(t)+\boldsymbol{D} \cdot e^{b l} \cdot \cos (2 \pi l) \tag{12}
X(t+1)=ω⋅Xp(t)+D⋅ebl⋅cos(2πl)(12)
随机差分变异策略如下:
X
(
t
+
1
)
=
r
1
×
(
X
p
(
t
)
−
X
(
t
)
)
+
r
2
×
(
X
′
(
t
)
−
X
(
t
)
)
(13)
\boldsymbol{X}(t+1)=r_{1} \times\left(\boldsymbol{X}_{p}(t)-\boldsymbol{X}(t)\right)+r_{2} \times\left(\boldsymbol{X}^{\prime}(t)-\boldsymbol{X}(t)\right) \tag{13}
X(t+1)=r1×(Xp(t)−X(t))+r2×(X′(t)−X(t))(13) 其中:
r
1
r_{1}
r1 和
r
2
r_{2}
r2 为
[
0
,
1
]
[0,1]
[0,1] 的随机数,
X
′
(
t
)
X^{\prime}(t)
X′(t) 为种群中随机选取的个体。 每个个体都要经过包围捕食、螺旋更新、搜索猎物阶段, 当 个体进行包围捕食或螺旋更新时采用自适应权重策略去更新位置,之后个体需要通过随机差分变异策略对其再次更新,取其变化前后的最优位置,加快了种群的收敛,有效地防止了种群陷入局部最优。种群通过这两种策略协同工作,使得算法具有更好的寻优效果。